Year: 2016
-

Iron reduction in soils of the Alaskan Arctic
From 2009-2012, we were funded by the NSF to study the importance of iron (Fe(III)) reduction as a dominant anaerobic process in soils of the Arctic Coastal Plain, near Barrow, Alaska. Fe(III) can serve as an alternative electron acceptor for anaerobic respiration when oxygen is depleted. What is special about Fe(III) is that it generally…
-
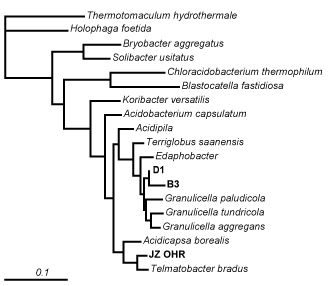
We’ve got Acidobacteria
Acidobacteria are highly abundant in most soil bacterial communities, but are rarely cultured in the lab. The ones that have been cultured grow very slowly and generally require specialized growth media. Acidobacteria probably represent the ultimate K-selected microbe: they very gradually build up populations and then hang on forever, dividing once a month or so…
-

Rate-yield tradeoffs and soil respiration in a high elevation forest
While studying seasonal changes in microbial growth kinetics in soils of a subalpine forest in the Colorado Rocky Mountains, I noticed a negative relationship between maximum growth rate and growth efficiency, or yield (the fraction of carbon consumed that is converted into biomass, as opposed to being respired as CO2). Turns out this is a…
-

Differential responses of native and exotic coastal sage scrub plant species to N additions and the soil microbial community
JDPE student, Francis Bozzolo, published part of his dissertation in Plant and Soil. Part of the motivation for this study was that the dominant shrubs of Coastal Sage Scrub ecosystems are generally clumped in patches, as in the photo above. Do they partition N by depth or chemical form? What happens to N partitioning when…